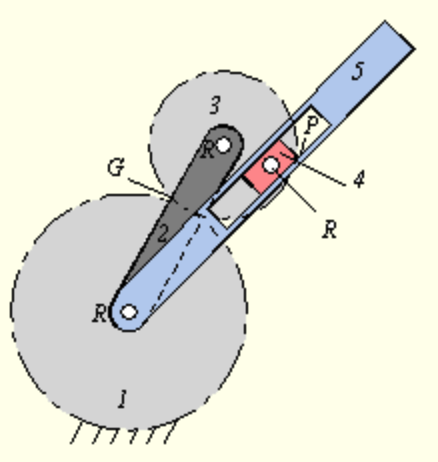
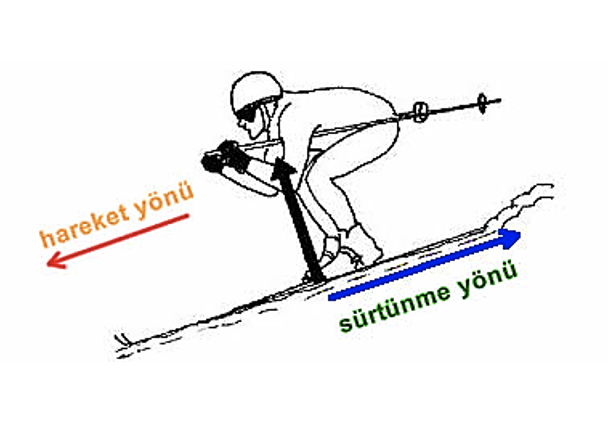
A free and open educational resource for faculty, students, and self-learners throughout the world. METU OCW does not grant credits or degrees and does not provide access to faculty. METU OCW gives you open access to the materials used in a variety of courses. Currently, 192 open educational resources are available.
To develop OpenCourseWare content and share existing materials, you just need to contact us via our email odtuzem@metu.edu.tr. The rest of the process, digitalizing your materials and uploading digital materials to the system, will be done by us. In addition, if you wish, we can record your lectures and add the content of the course.
METU OCW is awarded to: -- Open Education For Excellence Outstanding Site Award 2016 -- TBD Stars of Informatics 2013 E-Education Category 1st Prize -- Open Education For Excellence Multimedia Course 2011 -- TBD Informatics 2011 Best Educational Site Award